The
fossil record is life’s evolutionary epic that
unfolded over four billion years as environmental
conditions and genetic potential interacted in accordance
with natural selection. It could be likened to a movie
recording the history of life across nearly four billion
years of geological time. The problem is that only
a small fraction of the frames are preserved, and
those that have been preserved have often been chronologically
scrambled. Viewed in this way, reconstituting the
movie seems intractable, and yet science has done
so. Frames are still missing, but the intricate, interwoven
plots are largely revealed. The movie shows a cycling
planet, with life originating in the sea, and the
appearance and disappearance of, as Darwin
described in Origin
of Species in 1859, “endless forms most
beautiful and most wonderful”. The process of
descent with modification through natural selection,
or evolution, that Darwin
published in the mid 19th century became the framework
for many scientific disciplines to use fossils to
recover the missing frames and their ordering in time.
Some one hundred years after Darwin the mechanisms
of genetic memory and common descent have been elucidated
at the molecular level. In this new era of sequencing
the genomes and proteomes of multiple species, where
science can infer protein lineage dating back into
the Proterozoic, the tangible evidence in the imperfect
fossil record remains paramount to the delineation
of still missing frames of the movie. The
fossil record is life’s evolutionary epic that
unfolded over four billion years as environmental
conditions and genetic potential interacted in accordance
with natural selection. It could be likened to a movie
recording the history of life across nearly four billion
years of geological time. The problem is that only
a small fraction of the frames are preserved, and
those that have been preserved have often been chronologically
scrambled. Viewed in this way, reconstituting the
movie seems intractable, and yet science has done
so. Frames are still missing, but the intricate, interwoven
plots are largely revealed. The movie shows a cycling
planet, with life originating in the sea, and the
appearance and disappearance of, as Darwin
described in Origin
of Species in 1859, “endless forms most
beautiful and most wonderful”. The process of
descent with modification through natural selection,
or evolution, that Darwin
published in the mid 19th century became the framework
for many scientific disciplines to use fossils to
recover the missing frames and their ordering in time.
Some one hundred years after Darwin the mechanisms
of genetic memory and common descent have been elucidated
at the molecular level. In this new era of sequencing
the genomes and proteomes of multiple species, where
science can infer protein lineage dating back into
the Proterozoic, the tangible evidence in the imperfect
fossil record remains paramount to the delineation
of still missing frames of the movie.
Darwin
envisioned an incremental
graduation
in descent with modification to new forms for which
most frames of movie are non existent in the fossil
record. The movie frames that survive reveal great
leaps from apparent ancestor to descendent with no
intermediaries,
much a continuity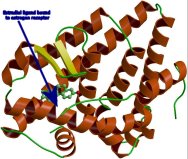 of incremental forms. Species or entire taxa suddenly
appear in the sparse fossil with no certitude of ancestry.
Others disappear with similar suddenness. Still other
fossils are enigmatic regarding ancestors and descendents,
and to which twig or branch of the great tree of life
they belong. The sometimes huge discontinuities in
the
fossil record and absence of transitional
forms worried
Darwin, and still provides the most demonstrable
evidence that creationists and proponents of intelligent
design proponents present to promulgate superstition
over science. Even without fossils, however, comparison
of sequences (of DNA, RNA and proteins) alone and molecular
phylogenetics analysis confirm evolution and deny
creationist
concepts. The process of evolution by natural selection
saves (i.e., conserves) important DNA coding sequences
over vast expanses of geological time. Molecular analyses
show that the proteins responsible for basic cell
organization
and function have truly ancient Precambrian origins.
A highly conserved sequence is a strand of DNA in
a
gene that contains a sequence of nucleotides that is
highly similar (i.e., homologous) across a wide range
of organisms. Such highly conserved sequences code
for proteins essential for survival, and have their
origins
in a common ancestor. Natural selection retains the
sequence since adverse mutations would likely impair
survival or reproductive function in the species. Among
the most commonly conserved genome sequences are
those
that code for the sites where small molecules bind
to protein receptors, often initiating gene transcription
of RNA, and subsequent protein production. For example,
the figure to the right depicts the primary estrogen
estradial (the small green molecule) attached to the
binding domain of the estrogen protein nuclear receptor
(the red ribbon). The estrogen
receptor is found to
be highly conserved in all extant vertebrate organisms
and is essentially to endocrine system functioning
in
reproduction, development, and throughout life. A significant
body of research supports origin of the nuclear receptor
protein into the precambrian. Research has demonstrated
that vertebrate estrogen receptors (even the ancient
lamprey are estradiol
(female sex hormone) receptors,
though non-vertebrate estrogen receptors (e.g.,
amphioxus or lancelet, marine organisms that may be
the most basal subphylum of chordates) are not; Ockham's
razor then suggests that most primitive estrogen receptors
were not able to bind estradiol but had a different
function, and that hormone regulation by estradiol
evolved in the vertebrate
lineage, possibly following gene duplication.
of incremental forms. Species or entire taxa suddenly
appear in the sparse fossil with no certitude of ancestry.
Others disappear with similar suddenness. Still other
fossils are enigmatic regarding ancestors and descendents,
and to which twig or branch of the great tree of life
they belong. The sometimes huge discontinuities in
the
fossil record and absence of transitional
forms worried
Darwin, and still provides the most demonstrable
evidence that creationists and proponents of intelligent
design proponents present to promulgate superstition
over science. Even without fossils, however, comparison
of sequences (of DNA, RNA and proteins) alone and molecular
phylogenetics analysis confirm evolution and deny
creationist
concepts. The process of evolution by natural selection
saves (i.e., conserves) important DNA coding sequences
over vast expanses of geological time. Molecular analyses
show that the proteins responsible for basic cell
organization
and function have truly ancient Precambrian origins.
A highly conserved sequence is a strand of DNA in
a
gene that contains a sequence of nucleotides that is
highly similar (i.e., homologous) across a wide range
of organisms. Such highly conserved sequences code
for proteins essential for survival, and have their
origins
in a common ancestor. Natural selection retains the
sequence since adverse mutations would likely impair
survival or reproductive function in the species. Among
the most commonly conserved genome sequences are
those
that code for the sites where small molecules bind
to protein receptors, often initiating gene transcription
of RNA, and subsequent protein production. For example,
the figure to the right depicts the primary estrogen
estradial (the small green molecule) attached to the
binding domain of the estrogen protein nuclear receptor
(the red ribbon). The estrogen
receptor is found to
be highly conserved in all extant vertebrate organisms
and is essentially to endocrine system functioning
in
reproduction, development, and throughout life. A significant
body of research supports origin of the nuclear receptor
protein into the precambrian. Research has demonstrated
that vertebrate estrogen receptors (even the ancient
lamprey are estradiol
(female sex hormone) receptors,
though non-vertebrate estrogen receptors (e.g.,
amphioxus or lancelet, marine organisms that may be
the most basal subphylum of chordates) are not; Ockham's
razor then suggests that most primitive estrogen receptors
were not able to bind estradiol but had a different
function, and that hormone regulation by estradiol
evolved in the vertebrate
lineage, possibly following gene duplication.
So
why have we digressed so far from the physical, macroscopic
world of fossils to the molecular world. There are two
reasons:
-
First,
to emphasize the astonishing accomplishments of Charles
Darwin (and many of his contemporaries) in using empirical,
phenotypical evidence (i.e, physical appearances)
from living and fossil organisms to correctly deduce
many aspects of how evolution works.
-
Second,
to affirm that the fossil record, or more particularly
its shortcomings and gaps about which Dawin worried,
is of much diminished concern due to modern molecular
phylogenetics across genotypes, that even without
fossils can infer decent with modification, and validate
theories of evolution.
The
fossil record, nonetheless, remains crucial to unfolding
major and minor branching points of the great tree of
life. Without fossils, inferring when genotype change
occurred requires the use of the molecular clock hypothesis,
which states that nucleotide or amino acid substitutions
(mutations) occur at a constant rate. In other words,
the amount of difference between two sequences can be
used to infer when in geological time that ancestral sequence
diverged occurred, thereby forming a phylogenetic branching
point. Actually, the rate of mutation differs among groups
of organisms, among genes, and even among different portions
of the same gene. Because of this, molecular clocks used
in molecular phylogenetics must be calibrated with fossils
to ascertain when groups and clades of organisms appeared.
To repeat, the fossil record is absolutely critical for
calibration in phylogenetics.
Creation
of the Fossil Record
First,
a working definition of the fossil record is needed.
We will use an overarching definition: all fossils known
to science. The question constantly arises: is the fossil
record complete? The short answer is, like the film
metaphor above, way far from it. The other question
is: Is the fossil record adequate? Here the answer is,
it depends on who you ask. A survey of the opinions
leads to the conclusion that the fossil record is pretty
much adequate, and constantly improving particularly
in recent decades. We will revisit these questions later,
but first a large amount of background is required.
It
is important to address the rarity of fossils in the
context that for any particular organism that once existed,
the probability that it today is part of the fossil
record is infinitesimally small. Such profound rarity
is a consequence of three factors: 1) fossil formation
is a rare event; 2) fossil survival is a rare event;
and 3) an exceedingly tiny fraction of surviving fossils
will ever be accessible to be found, though the crust
of the earth is filled with them. We’ll examine
these factors in sequence.
Fossil
Formation
The
likelihood of an organism becoming fossilized is poor,
and even less likely is that more than a small portion
of tissues will become fossilized. If death is the primary
prerequisite for fossil formation, then the second prerequisite
is the forestalling of the gory ravages of decay, where
the deceased becomes an ecosystem and feast unto its
own for an astonishing diversity of life forms from
microscopic bacteria to large predators. The transformation
of an individual deceased organism into a fossil is
a probabilistic rarity, and the vast majority of remains
that are not eaten are either turned to dust or dispersed
as dissolved molecules.
The
science of fossil formation is complex in terms of biological,
chemical, and physical processes, and is multidisciplinary,
but many aspects are part of a relatively new field
termed Taphonomy, which studies the decay and destruction
of an organism’s remains through time.
We
can envision that, as the ancient earth’s crust
cooled, a time was eventually reached when rain from
a dense atmosphere containing all the water was not
immediately evaporated. Once begun, hard rains fell
and fell for countless millennia finally filling the
awaiting ocean basins. With the first rainfall, the
inexorable processes of erosion and dissolution began,
wearing away the then barren earth’s crust and
ultimately filling the awaiting ocean basins with not
only water, but also the dissolved elements needed for
life in its most primitive manifestation to appear in
the primeval seas. 
 The
rain begat the seas, where life appeared, and began
wearing the land away, forming the sediments needed
to cover and preserve the traces of living organisms
known as fossils. The formation and transport of sediments
might be likened to a never ending snow storm. Relentlessly
the sediments have been carried by the rainwater, down,
down, down by the inexorable force of gravity. Perhaps
the flakes were sometimes light and sparse, and other
times a veritable blizzard, but without cessation. The
sediments were also an implacable force grinding away
everything in its path. We now know that geological
time is replete with orogeny (Greek for "mountain
generating") events that are the result of plate
tectonics. These events have pushed up huge mountain
ranges, on the land, or in the seas, many of which have
been mostly or completely worn away (see example –
the Appalachian Orogeny). Mountains that were miles
high have been ground down to remnants of former greatness,
testimony to the scale and power of the rain and the
sediments. The
rain begat the seas, where life appeared, and began
wearing the land away, forming the sediments needed
to cover and preserve the traces of living organisms
known as fossils. The formation and transport of sediments
might be likened to a never ending snow storm. Relentlessly
the sediments have been carried by the rainwater, down,
down, down by the inexorable force of gravity. Perhaps
the flakes were sometimes light and sparse, and other
times a veritable blizzard, but without cessation. The
sediments were also an implacable force grinding away
everything in its path. We now know that geological
time is replete with orogeny (Greek for "mountain
generating") events that are the result of plate
tectonics. These events have pushed up huge mountain
ranges, on the land, or in the seas, many of which have
been mostly or completely worn away (see example –
the Appalachian Orogeny). Mountains that were miles
high have been ground down to remnants of former greatness,
testimony to the scale and power of the rain and the
sediments.
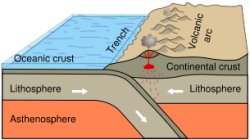
A)
Oceanic-continental convergence resulting in subduction
and volcanic arcs illustrates one effect of plate
techtonics.
Although they are present, transitional forms are
rare in the fossil record. In fact, the fossil record
demonstrates that some species have remained essentially
unchanged for millions of years. This makes sense,
because if a species is well adapted to its environment,
its current traits will continue to be selected for.
A species will only undergo major change, if its environment
changes in a way which leaves it significantly less
well adapted to survive. Such transitions tend to
occur rapidly. A mammal species for example, might
evolve into another distinct mammal species in less
than a hundred thousand years. In the history of life,
that is a short period of time. Speciation of plants
and simpler animals, can occur much more rapidly.
This evolutionary pattern of long periods of geologic
time with little change, punctuated by short periods
of rapid change, is referred to as punctuated equilibrium.
|