StromatolitesThe Oldest Fossils |
|
Related
Pages: | ||||
The Oldest FossilsStromatolites
are not only Earth's oldest of fossils, but are intriguing
in that they are our singular
visual portal (except for phylogenetic
determination of conserved nucleic acid sequences and some
subtle molecular fossils) into deep time on earth, the emergence
of
life,
and the evolving of the beautiful forms of life of modern time.
A small piece of stromatolites encodes biological activity
perhaps
spanning thousands of years. In broad terms, stromatolites are
fossil evidence of the prokaryotic life that remains today,
as it has always
been, the preponderance of biomass in the biosphere. For those
that subscribe Scientists disagree on how to define stromatolites. A common definition goes something like: A lamiated rock formed by the growth of blue-green algae (i.e., cyanobacteria)". This definition is, in fact, such a gross oversimplification as be scientifically useless. It does contain a modicum of truth, however, in that the largest volume of stromatolitic formations was likely formed by biogenic processes involving photosynthetic cyanobacteria. Cyanobacteria’s metabolic byproduct, oxygen, rusted the earth, pumped enormous oxygen poison to them into earth’s atmosphere, and in so doing paved the way for aerobic-based life to emerge and diversify; cyanobacteria’s contributions to life led to their own prodigious decline.
Excluding some exceedingly rare Precambrian fossils such as the Russian White Sea Ediacaran fauna, stromatolites and thare the only fossils encoding the first 7/8th of the history of life on earth. They encode the role that ancient microorganisms played in the evolution of life on earth and in shaping earth's environments. The fossil record of stromatolites is astonishingly extensive, spanning some four billion years of geological history with the forming organisms possibly having occupied every conceivable environment that ever existed on earth. Today, stromatolites are nearly extinct in marine environments, living a precarious existence in only a few localities worldwide. Modern stromatolites were first discovered in Shark Bay, Australia in 1956, and throughout western Australia in both marine and non-marine environments. New stromatolite localities have continued to be discovered in various places such as the Bahamas, the Indian Ocean and Yellowstone National Park, to name but a few localities.
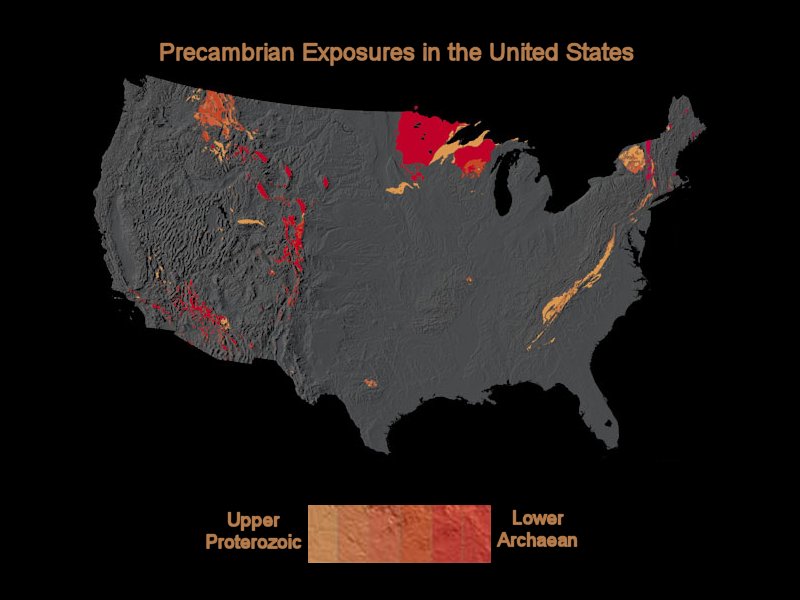
Stromatolite Stratigraphy:Stromatolites occur throughout the world, but become diminishingly uncommon in the Archaean. Earth’s violent surface, floating on tectonic plates, is subjected to volcanism, subduction (into the earth’s mantle), uplift, metamorphism, and enormous erosion forces. Probably many Archaean rocks have succumbed to these many forces. So too have many Stromatolites from the Proterozoic. Production of stromatolites has been relatively minor during the Phanerzoic Eon with its entirely different reef ecosystems. What rock has not been lost may of course be buried deeply. The figure to the left shows Precambrian exposure within the United States. The figure indicates the paucity of localities where finding the oldest fossils exists. The north, central U.S. and Wisconsin and Minnesota particularly have much old rock, and not surprisingly, considerable Stromatolites and banded iron.
As previously noted, stromatolites are most often described as biogenically-produced structures formed by colonies of photosynthesizing cyanobacteria. However, this is an enormous oversimplification given that the weight of scientific evidence suggests that all three domains of life (the Archaeans, Eubacteria, and Eukaryotes) appeared in the Archaean Era, and thus the so-called microbial mats would have contained representatives among all three domains. Just how and when the base of the tree of life split into the three main branches remains one of the most important questions in all of biology and science, and is the source of constant scientific dispute. Which of the prokaryotes came first, the Archaeans or the Eubacteria remains unresolved, and a consensus has emerged that these primitive microorganisms laterally exchanged genes further confounding attempts to validate what begat what during to course of early evolution on earth. Lateral gene transfer belies the concept of the single common ancestor (see Woese, 1998). While formation by colonies of cyanobacteria is probably the primary mechanism for formation of surviving stromatolites in the deep time of the Archaean and half way through the Proterozoic, it is unlikely to have been the only mechanism. Recent research (Gupta, 1998a, 1998b, and an extensive literature) indicates the other prokaryotic and the most genetically diverse domain of life, the Archaeans, evolved alongside each other and possibly swapped genes with the Eubacteria. All prokaryotes (both Eubacteria and the Archaeans reproduce by cell division (binary or multiple fission) and, lacking sex, are essentially clones and among the slowest evolving organisms. Moreover, some microfossils (actually, putative ancient cellular remnants) indicate that primitive Eukaryotic microorganisms may have appeared prior to 3.5 Bya. Thus, before the end of the Archaean time some 2.5 Ba, all three domains of life (Eubacteria, Archaea, and Eukaryotes) co-existed and were likely already quite diverse. Some were autotrophs, some chemotrophs and some heterotrophs, and collectively they had a multiplicity of metabolic processes from which to derive their energy, and as they do today. Just as microorganisms were extremely diverse in deep time, so were there a corresponding extreme diversity of biogenic and chemical (abiogenic) mechanisms that are plausible for the formation of laminar carbonate and other structures that we call stromatolites, and the possible ancient cellular microstructions they might contain. Ascribing all stromatolite formation in the Archaean and Proeterozoic to cyanobacteria, as is often seen in general literature, is incorrect. Whether or not stromatolites contain preserved cellular structures (microfossils) also remains highly contencious, especially in older Arachean rocks. Viewing these putative ancient cells requires special polishing techniques and high magnification. Additionally, Molecular fossils (or fingerprints) based on atom ratios in Archaean sediments is highly contencious, but scientifically critical, since such subtle molecular traces found in Australia has led to conjecture that microorganisms with nuclei appeared before 3.8 Ba. Milestones in Evolutionary Biology - Bacteria, Archaea and EukaryaStromatolites may hold the key to determining one of the most important questions in evolutionary biology, how and when the tree of life branched into the three domains, the Bacteria, Archaea and Eukarya. Of particular importance is when microorganisms with advanced photosynthetic metabolism first appeared, since molecular phylogenetics suggests that all three domains would have already appeared and significant evolution must have already taken place (Schopf, 2000 and Olson 2006). Equivalently important is when the eukaryotic nuclear genome became a chimera with contributions from both the Archaean and Bacteria (Gupta, 1997). While there is scientific consensus that phosynthetic cyanobacteria became prodigious at some point in the Archaean, as evidenced by first the rusting of the earth and ultimately the oxygenation of the atmosphere, just when that occurred remains both uncertain and contentious. More generally, stromatolitic laminate structures themselves as well as the putative cellular structures and molecular signatures they sometimes contain have been and remain controversial, since they can be explained by either biogenic or abiotic processes (Grotzinger, 1999). Further confounding the definition of what is or nor not stromatolites is known processes for bacterially mediated precipitation of minerals (Paerl, 2001). Warrawoona Group in Western Australia - a scientific dispute The putative stromatolites with microstructures resembling bacteria from the extensive stromatolitic formations of the 3,430-million-year-old Strelley Pool Chert within the Warrawoona Group in Western Australia have been hotly debated ever since their discovery by Lowe (1980, 1983). Lowe (1994) later ascribed conical form genera to abiotic evaporative precipitation, as did Grotzinger (1999), and Brasier (2002) also found no support for the microfossils as biomarkers. Whether microstructures within the Warrawoona Group stromatolites are the imprints of ancient filamentous and possibly photosynthetic microbes as argued by Schopf (1987, 1993) and Awramik (1992) became a heated debate that remains unresolved. A recent and extensive study of seven distinct stromatolitic form genera by Allwood (2006) certainly lends support to proponents of biogenetic origins of the chert, since the simultaneous set of forms is more difficult to explain with known abiogenic processes. However, whether the microstructures are fossil microbes remains unresolved. If they are microbe fossils, there would still remain the critical question of what type they are, archaea, cyanobacteria, another type of photosynthetic bacteria, chemosynthetic bacteria, or some combination of these. Gunflint Chert - abundant preserved cells
Schoff and coworkers report unambiguous cyanbacteria microfossils from the Archaean, 2.6 Bya Campbell Group in Cape Province, South Africa (Altermann, 1995), and numerous Proterozoic sites, including the stromatolitic black chert from the approximately 850 Mya Bitter Springs Formation, Australia (House, 2000), the 680-790 Mya Min'yar Formation of the southern Ural Mountains (Nyberg, 1984). Recently, Schopf, perhaps the most outspoken proponent of the evidence of cyanobacteria in the early Arachaean, listed 48 Archaean deposits reported possibly containing biogenic stromatolites, of which 14 contain 40 morphotypes of putative microfossils, and 13 are in the age range of 3.2 to 3.5 Bya (Schopf, 2006). Other researchers strongly disagree. For example, Blank (2002), based on genome sequencing, posits that cyanobacteria may have originated as late as 2.3 billion years ago. The phylogenetic tree based on whole genomic DNA sequences show that cyanobacteria were one of the last major lineages to diverge off the bacterial tree, and were preceded by sulfur-oxidizing bacteria and sulfate-reducing bacteria. This mirrors the changes in the geochemical record, centered around 2.7 billion years ago. The hypothesis is consistent with geology that finds isotopic fractionation of sulfur compounds becomes large, followed by the sudden increase in oxygen in the atmosphere and surface water environments at about 2.2 or 2.3 Ba. Oxygenation of the Atmosphere - a profound transformation of the bioshere Regardless of when the cyanobacteria appeared, it is widely accepted that they comprised the predominant form of life on early earth for some two billion years, and were responsible for the creation of earth's atmospheric oxygen, consuming CO2 and releasing O2 by photosynthetic metabolism. Creation of the modern atmosphere is, of course, perhaps the most critical event in geological history that powered the Cambrian Explosion and subsequent evolution of the aerobic forms of life, including all animals (Ohno, 1997). Microbial Mats to Stromatolitic Structures
Stromatolites are also variously described as being formed by algae that are, in turn, assumed to be plants; this description still persists in old textbooks and on the Internet, but is vary incorrect. It is a holdover from a time that cyanobacteria were thought to be algae (and were called blue-green algae) and from when algae were thought to be plants. Actually, cyanobacteria are prokaryotic bacteria (domain of life Eubacteria), and "genomic" science is sill debating whether eukaryotic, photosynthetic, and autotrophic algae are plants or deserve a distinctive phylogenetic grouping. Regardless, the eukaryotic algae did not appear until about 1.5 Ba, some 2 billion years after stromatolites significantly began forming. It is therefore likely that stromatolite formation by algae was not significant until the Phanerozoic, or possibly the Late Proterozoic.
Recent research supports the hypothesis that stromatolite form diversity increased through the Paleoproterozoic, reached a maximum in the Mesaproterozoic at about 1.5 Bya that persisted to about 700 Ma, and steadily declined to several taxa by the Precambrian-Cambrian boundary (Olcott, et. al., and Wray, 1996). This is in contrast to previous data indicating a steep decline at 2 Bya that now appears from the data to be an artifact of 50% of all stromatolites coming from a single basin and author; thus, this decline was in volume. When this regional anomaly is removed, the steepest decline in forms appears to have occurred in the Cambrian. By normalizing stromatolite forms with volume of preserved carbonate rock, the authors posit that the steepest decline in stromatolite form diversity occured in the late Neoproterozoic, and culminated in the Lower Cambrian, coincident with the widespread appearance of macroscopic metazoa and significant bioturbation (i.e., activity of bottom-living animals that keeps sediments oxygenated and homogenous). Conversely, stromatolites indirectly support the hypothesis that the diversification of major animal phyla occurred between 1 and 1.2 Ba (Wray, 1996). Since laminated sediments are a sign of oxygen depletion in the bottom zone of the sea, bioturbation would inhibit the building of stromatolitic structures. This research seems consistent with evolutionary theory that would anticipate diversification of stromatolite forming taxa due to selective pressure from other organisms that were emerging and themselves diversifying. We will likely have no more than a sketchy understanding of the paleoenvironments in which stromatolites were formed in the deep Precambrian time, and only an incomplete understanding of the environments in the Paleozoic. Sound conjecture is possible if we examine the now rare environments that support stromatolitic growth during modern times. Cyanobacteria are found to be a primary organism in the formation of modern microbial carbonates. These prokaryotic bacteria (slang name is blue-green algae owning to pigmentation involved in photosynthesis) are now only found in areas where there is reduced grazing and burrowing by other organisms, and a low occurrence of macro-algae and plants. Environments where modern stromatolites are found typically are hypersaline, but also include areas of high alkalinity, low nutrients, high or low temperatures, and strong wave or current actions. The obvious pattern emerges that modern stromatolites tend to exist in areas that most other life forms consider less desirable or possibly intolerable. Thus, organisms producing modern stromatolites are generally limited to areas where organisms with which they have to compete and/or organisms that might use them for nutrients are not prevalent. Also see: Stromatolite References Evolution of Multicellularity Endosymbiosis Leads to Mitochondria Endosymbiosis Leads to Chloroplasts Museum Cambrian Explosion Fossils | ||||||







|
| ||||||
Fossil
Museum Navigation:
Home Geological Time Paleobiology Geological History Tree of Life Fossil Sites Fossils Evolution Fossil Record Museum Fossils |